
tRNA-Isodekoder: RNA-Moleküle mit unerwarteter regulatorischer Funktion
Genetik ist komplizierter als das Schulbuch behauptet

In der Schule muss man didaktisch reduzieren. Das Problem ist, wenn diese didaktische Reduktion an der Uni oder später nicht aufgeklärt wird, damit wird sie zur Lüge, obwohl das Wissen generell vorhanden wäre.
Ich bin immer wieder entsetzt über die Wissenslücken bei den genetischen Grundlagen, die mir selbst unter Wissenschaftlern begegnen. Viele Wissenschaftler sind immer noch an ihrem reduktionistischen, simplistischen Schulweltbild, wie mRNA in Protein übersetzt wird, verhaftet. Dann passieren natürlich solche “Unfälle” wie modRNA-Injektionen.
Wenn man keine Ahnung von der Komplexität des Systems hat und immer noch auf dem Niveau eines Schülerdudens agiert, bringt man unter Umständen viele Menschen um.
Was ist tRNA und wie sieht sie aus?
Was tRNA ist, lernt man in der Schule. Das ist die RNA, welche dem Ribosom hilft, mRNA in Proteine zu übersetzen.

Im Schülerduden sieht das so aus, das ist definitiv Schulstoff.

tRNA ist selbst eine Form der RNA, die im 3D-Raum komplex gefaltet ist. tRNA ist eine der wenigen RNAs von denen wir überhaupt Kristallstrukturen haben, daher wird an tRNA die Primär, -Sekundär und Tertiärstruktur von RNA gelehrt. Also nein, RNA ist kein einfacher Strang, RNA ist auch kein Strang der schön gefaltet ist wie Struktur a) im nächsten Bild.
Der Unterschied zwischen sekundär a) und tertiär b) Struktur. Das ist Lehrbuchwissen, das ich schon 1997 in der Vorlesung gelernt habe. Das ist nicht neu, das ist keine Raketenwissenschaft. RNA ist im 3D-Raum auf eine Art gefaltet, die wir aktuell weder verstehen noch vorhersagen können, weil es kaum Kristallstrukturen gibt, an denen man lernen könnte.
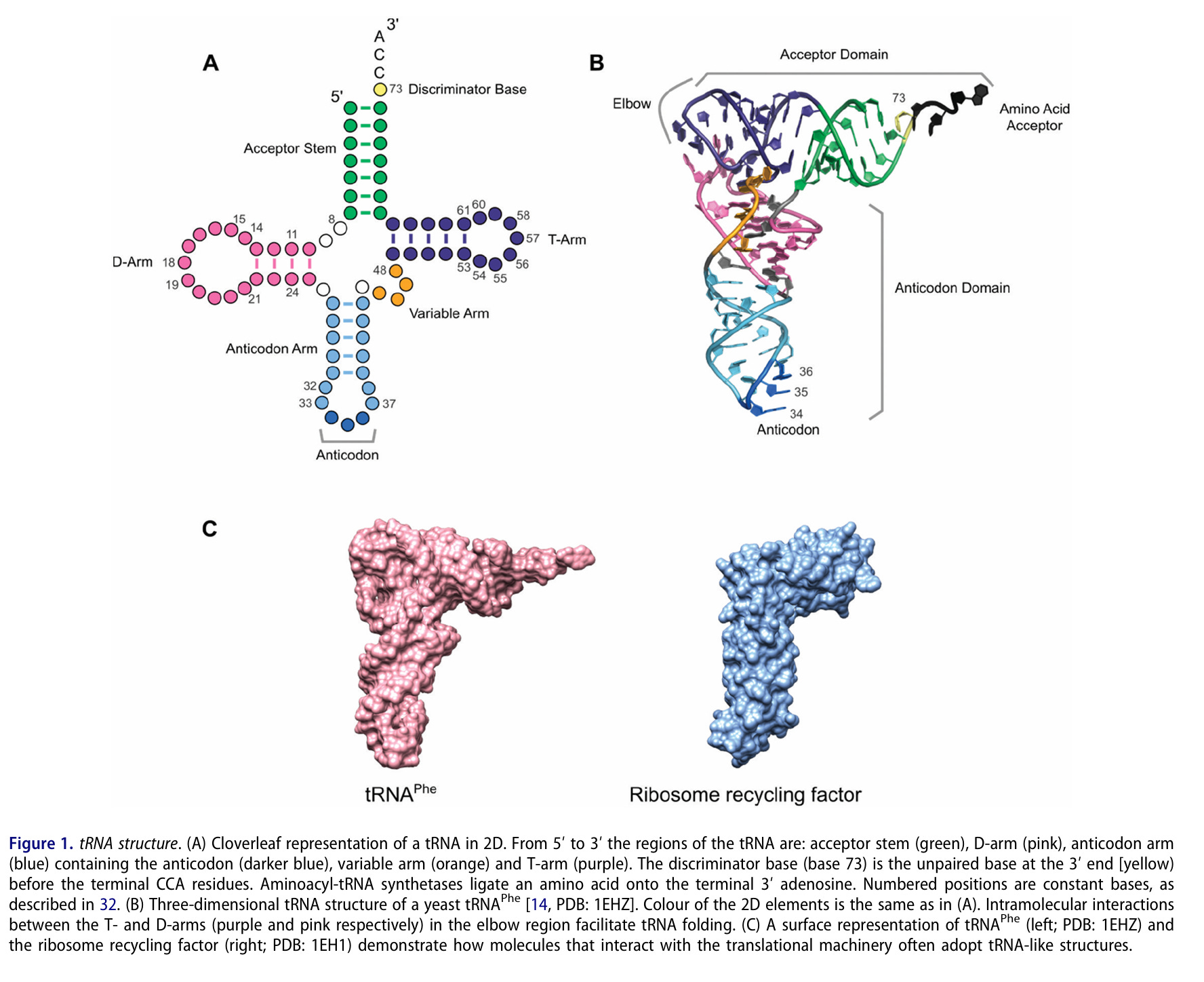
So sieht tRNA wahrscheinlich in 3D aus:
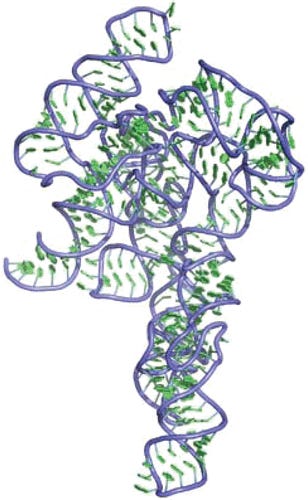
Der genetische Code wird nicht dekodiert, wie man es in der Schule lernt.
In der Schule lernt man, dass es einen Triplet Code gibt, und jedes Triplet eine tRNA hat und so mRNA in Proteine dekodiert wird.

Das ist leider falsch und deutlich komplexer und teilweise leider immer noch ungeklärt.
Nicht jedes Codon hat überhaupt eine tRNA. Es gibt Codons, die haben gar keine eigene tRNA und werden von anderen tRNAs mit bedient über nicht watson-Crick Paarungen (wobble).

6 Rot markiert die Codons, die keine eigene tRNA haben und nur über sogenannte wobbel tRNA (also an einer Stelle rumwackelnde tRNA bedient) werden, andere Codons von tRNAs wo sowohl als auch zutrifft. Was passiert wohl, wenn da ein N1-Methylpseudouridin verbaut ist?!

“RNA wird durch zelluläre RNasen abgebaut und dem Nukleinsäurestoffwechsel unterworfen. Der Nukleotidstoffwechsel findet in der Zelle kontinuierlich statt, wobei das Nukleosid zu Abfallprodukten abgebaut und ausgeschieden oder für die Nukleotidsynthese wiederverwendet wird. Daher werden keine Studien zum RNA- oder Proteinstoffwechsel oder zur Ausscheidung durchgeführt.”7
Die Dekodierung variiert vermutlich leicht von Organismus zu Organismus und von Organ zu Organ. Hinweise darauf gibt es von der Seidenraupe bereits seit 1979.8 Man kann das auch in Zellkultur beobachten, dass verschiedene Zellkulturlinien/Zellarten ein Protein durchaus unterschiedlich herstellen, selbst bei Bakterien. Es hat seinen Grund, warum es für Proteinproduktion extra Expressionsstämme gibt und nicht jeder Stamm für jedes Protein geeignet ist.
Schon die Mitochondrien dekodieren einige Codons abweichend vom Zellkern.9 10 11 12
Anders als durch diese alternativen Codon Dekodierungen ist nicht zu erklären, warum wir so viele verschiedene Proteine haben, bei so wenigen Genen. Das ist alles nicht neu. Das habe ich schon in den späten 1990er Jahren an er Uni gelernt. Ich wiederhole hier teilweise nur Stoff aus der Grundstudiumsvorlesung “Einführung in die Genetik” aus dem ersten Semester 1997.
Für tabellarische Darstellungen von wissenschaftlichen Publikationen ist die google KI durchaus nützlich. Hier die unterschiedliche RNA Dekodierung von Zellkern und Mitochondrium:

Eine Wobble-Bindung (auch Wackelbasen-Paarung genannt) ist eine nicht-standardisierte Basenpaarung bei der Proteinbiosynthese, die von den klassischen Watson-Crick-Regeln (A-U, G-C) abweicht. Sie beschreibt die chemische Eigenschaft, dass die dritte Base eines mRNA-Codons lockerer gebunden ist und sich flexibel mit verschiedenen Basen des tRNA-Anticodons paaren kann. 13

Abbildung 3. Codonhäufigkeiten beim Menschen. (A) Die Häufigkeit des Vorkommens von Aminosäuren (AA). Die Aminosäuren sind auf der Abszisse in Ein-Buchstaben-Notation aufgeführt. Die Häufigkeit, mit der die jeweilige Aminosäure kodiert wird, ist auf der Ordinate als Prozentsatz angegeben. (B) Normalisierte Häufigkeit des Vorkommens von Aminosäuren. Für jede Aminosäure wird die Aminosäurehäufigkeit (%) auf die Anzahl der synonymen Codons normiert. (C) Die Häufigkeit des Vorkommens von Codons. Die Codons sind auf der Abszisse des Balkendiagramms aufgeführt; jeweils ist die Verwendungshäufigkeit (in Prozent) auf der Ordinate dargestellt. Die Codons sind nach ihrer gemeldeten Häufigkeit geordnet. Stoppcodons sind nicht aufgeführt. Weiße Balken kennzeichnen Codons, die nur von entsprechenden kognitiven tRNAs decodiert werden können. Schwarze Balken kennzeichnen Codons, für die kein kognates tRNA-Gen vorhanden ist und die nur durch Wobble-tRNAs entschlüsselt werden können. Graue Balken kennzeichnen Codons, die sowohl durch kognitive als auch durch Wobble-tRNAs entschlüsselt werden können. Es ist zu beachten, dass potenzielle U-G- und I-A-Wobble-Wechselwirkungen in diesem Balkendiagramm nicht berücksichtigt werden, da diese Wobble-Wechselwirkungen beim Menschen offenbar noch nicht bestätigt wurden. (D) Normalisierte Codonhäufigkeit. Die Codonhäufigkeit (%) wurde normalisiert, indem die Codonhäufigkeit durch die durchschnittliche Anzahl der kognitiven und Wobble-tRNA-Gene für jedes Codon dividiert wurde. Die Daten zur Codonverwendung beim Menschen stammen aus der „Codon Usage Database“ (http://www.kazusa.or.jp/codon/).14
“Aufgrund des Wobble-Effekts reichen 32 tRNA-Spezies aus, um alle 61 Codons zu dekodieren. Pro Organismus gibt es jedoch viele verschiedene Isodekoder, von denen jeder oft mehrere genomische Kopien aufweist. Die zusätzlichen Kopien sorgen für Redundanz im Falle einer Funktionsverlustmutation und dienen als Puffer, falls eine Mutation zu einer nicht-kognaten Dekodierung führen sollte. Diese zahlreichen Kopien bilden das Ausgangsmaterial für die Evolution des genetischen Codes.”15
Könnte man den Verbau von N1-Methylpseudouridin (m1Ψ) unter “Evolution” des genetischen Codes verbuchen? Könnten Codons mit m1Ψ von tRNA-Isodekodern alternativ dekodiert werden? Hat man das untersucht oder auch nur in Betracht gezogen? Inwieweit könnten alternativen Proteine entstehen, die wir nicht finden, weil wir nicht wissen, wonach wir suchen müssen?
Was sind tRNA-Isodekoder?
tRNA Isodekoder hatte ich schon in meinen Buch und den Artikeln zu den grenzdebilen Schwachsinnsideen in Protein Engineering erwähnt. Wenn man nicht einmal weiß, wie mRNA zu Proteinen übersetzt wird, dann pfuscht man nicht in die Kodierung.
Die Anzahl der tRNA-Isodekoder hat bei Säugetieren drastisch zugenommen, doch die genauen molekularen und physiologischen Gründe für diese Zunahme sind nach wie vor unklar. […] tRNA-Isotypen sind mit einer der 20 Aminosäuren beladen und lassen sich in verschiedene Isoakzeptoren – tRNAs, die synonyme Codons für dieselbe Aminosäure verwenden – sowie in verschiedene Klassen von Isodekodern (Abb. 1a) einteilen, d. h. tRNAs, die dasselbe Codon verwenden, sich jedoch im tRNA-Körper unterscheiden, wobei jede von ihnen aus einem oder mehreren tRNA-Genen transkribiert werden kann.16
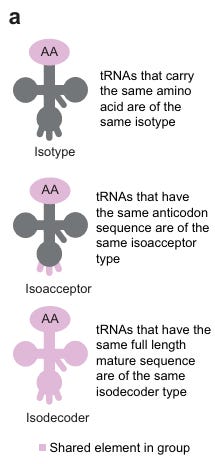
Es ist also noch komplexer als in der Einleitung erklärt. Es gibt nicht nur wobble und passende tRNAs, es gibt dazu noch tRNAs, die im Stamm unterschiedlich aussehen.
Die gesammelten Erkenntnisse belegen, wie wichtig die Vielfalt verschiedener Isoakzeptoren ist, da sie die Effizienz und Genauigkeit der Translation von mRNAs mit unterschiedlicher Codonverwendung bestimmen. Die Dominanz der RNA-Isodecoder – die mehr als die Hälfte aller bei höheren Säugetieren beobachteten RNA-Gene ausmachen – war jedoch unerwartet, und ihre funktionelle Bedeutung ist bislang noch nicht geklärt. Die bioinformatische Auswertung von Genomsequenzen auf tRNA-Gene ergab, dass die Anzahl der tRNA-Isodekoder stark variiert und bei verschiedenen Organismen zwischen 3,6 und 55 % der tRNAs liegt, wobei bei Mikroorganismen nur sehr wenige beobachtet wurden, bei mehrzelligen Organismen eine Zunahme festzustellen ist und die höchsten Zahlen bei Säugetieren zu verzeichnen sind.”17
“tRNA werden in der Regel in drei Kategorien unterteilt:
i) Isotypen – verschiedene tRNA, die dieselbe Aminosäure transportieren;
ii) Isoakzeptoren – verschiedene tRNA, die dieselbe Aminosäure transportieren, aber unterschiedliche Antikodonsequenzen aufweisen; und
iii) Isodekoder – verschiedene tRNA-Moleküle, die dieselbe Aminosäure transportieren und dieselbe Antikodonsequenz aufweisen, sich jedoch durch kleine Sequenzunterschiede im tRNA-„Körper“ (alle Nukleotide außer dem Antikodon) unterscheiden. Isodekoder sind am schwierigsten zu unterscheiden, da sie sich unter Umständen nur durch ein einziges Nukleotid unterscheiden.”18
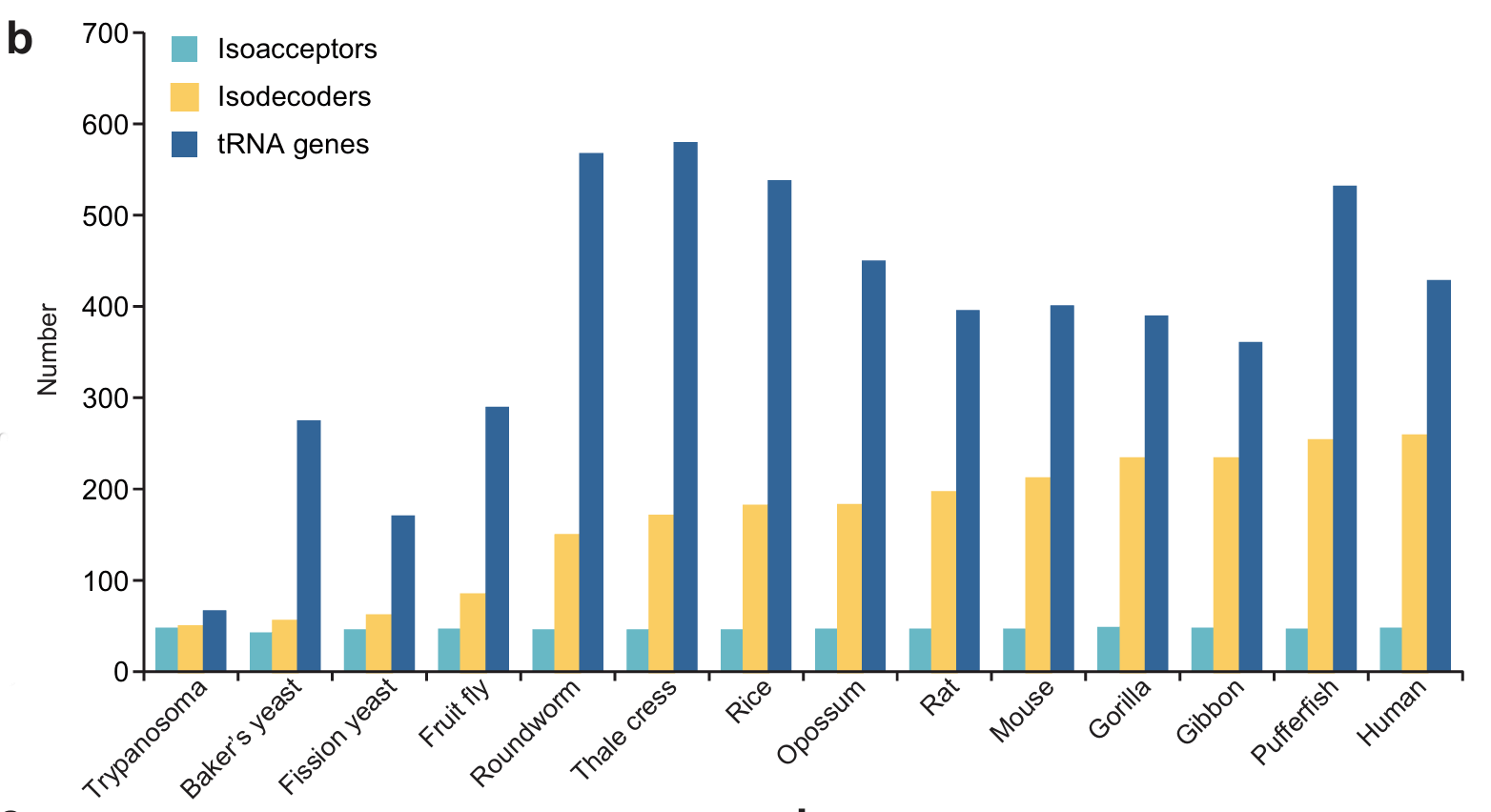
Es gibt deutlich mehr Isodekoder als Isoakzeptoren. Zur Erinnerung: Isoakzeptoren sind die tRNAs, die man in der Schule lernt.
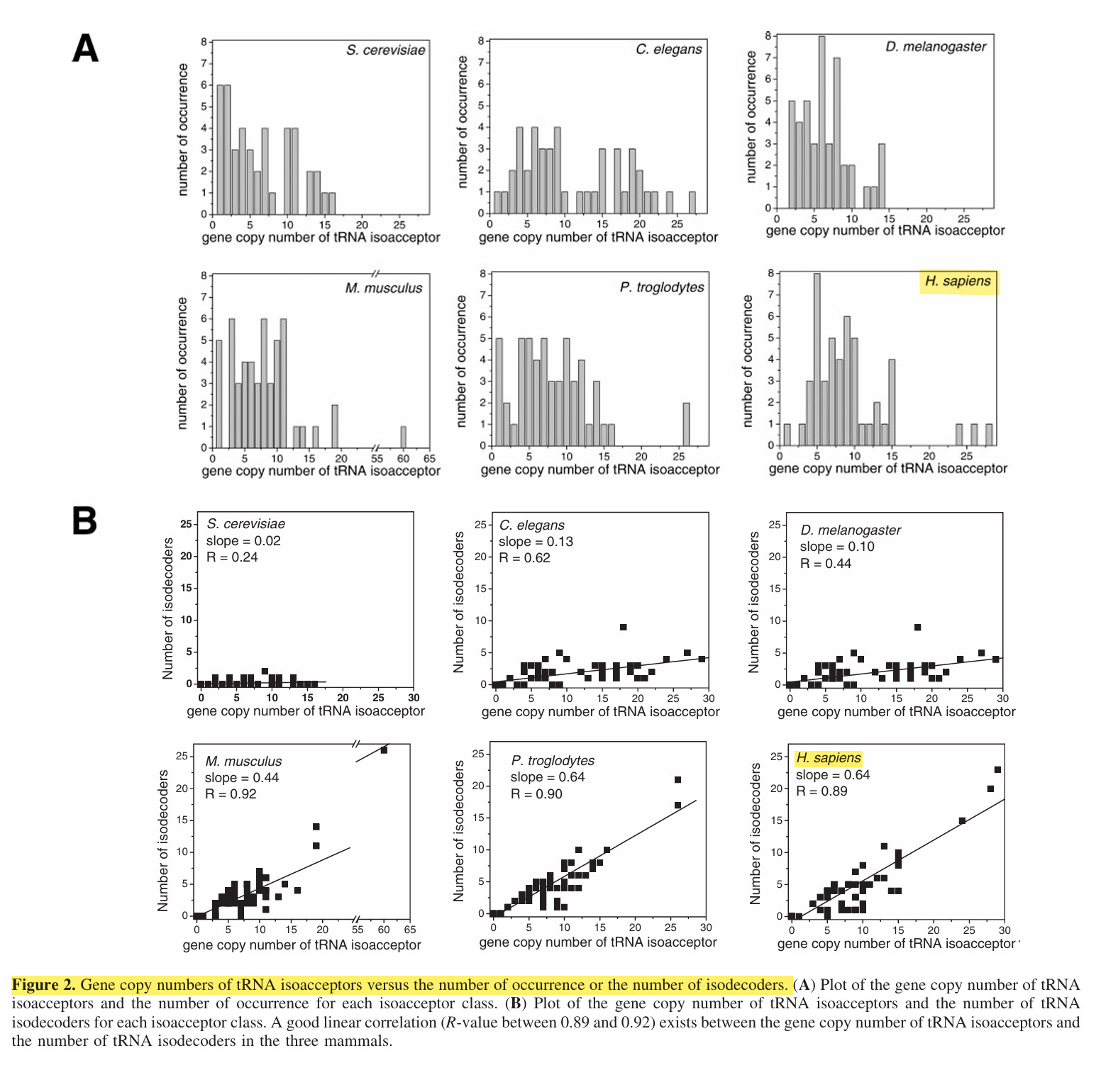
Die tRNA-Gene des Menschen weisen bei 6,4 % dieser variablen Nukleotide in Feld A und bei 12,3 % in Feld B Abweichungen auf. Diese Sequenzunterschiede können zu einer unterschiedlichen tRNA-Expression in menschlichen Geweben oder Entwicklungsstadien führen.20
Dennoch haben unterschiedliche evolutionäre Zwänge zur Entstehung verschiedener tRNA-Gene geführt, und In-vitro-Studien haben gezeigt, dass Unterschiede in den Sequenzen des tRNA-Körpers die Effizienz der Aminoacylierung und die Genauigkeit der Proteinsynthese beeinflussen können, was für bestimmte Prozesse von Nutzen sein könnte, beispielsweise zur Erleichterung der Proteinfaltung oder der Biogenese großer Proteinkomplexe.21
Hat man das bedacht bei den Plörren? Weiß das überhaupt jemand außer jenen, die daran forschen?
Die Rollen spezifischer Isodecoder-tRNA-Phe-Gene und die kompensatorischen Reaktionen auf deren Verlust variieren zwischen den Geweben, was die Gründe für die Expansion der Isodecoder-Gene bei Säugetieren verdeutlicht. […]
Wichtig ist, dass wir zeigen, dass innerhalb der tRNA-Phe-Familie die Expression von mindestens acht Allelen, einschließlich des spezifischen Isodecoder-Gens tRNA-Phe-1-1, für das Überleben und die frühe Embryonalentwicklung erforderlich ist. […] Mutationen in tRNAs, die vom mitochondrialen Genom kodiert werden, sind für den Großteil der mitochondrialen Erkrankungen verantwortlich und wurden bereits ausführlich untersucht. […]
Untersuchungen haben gezeigt, dass Unterschiede im tRNA-Körper deren Aminoacylierungs-Effizienz, ihre Wechselwirkungen mit dem Ribosom oder ihre Fähigkeit, Stoppcodons zu unterdrücken, beeinflussen können22
tRNAs, vor allem auch die Isodekoder, variieren in ihrem Vorkommen zwischen den Organen und sind wichtig bei der Embryonalentwicklung. Sie haben einen großen Einfluss darauf, wie Proteine produziert werden und falten und möglicherweise noch andere Funktionen.
N1-Methylpseudouridin (m1Ψ), welches in modRNA-Produkten verbaut ist, wurde 1976 im Bakterium Streptomyces platensis entdeckt23 und kommt in in der Natur direkt als Baustein in den tRNAs bestimmter Organismen vor.24 In höheren Zellen kommt m1Ψ in möglicherweise in ribosomalen RNA (rRNA) vor. 25
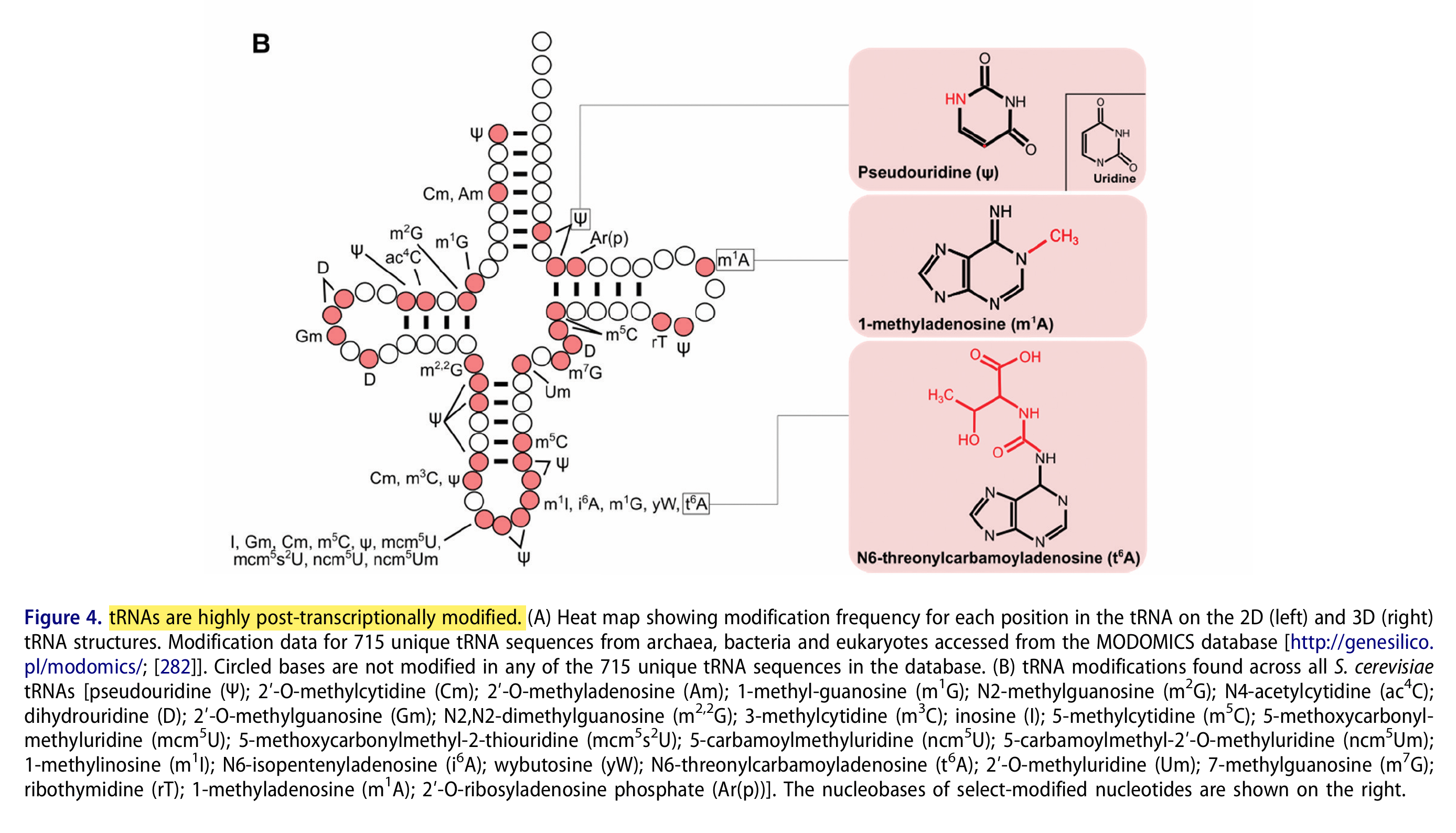
tRNAs werden in hohem Maße posttranskriptionell modifiziert.26 Was kann ein falsch verbautes m1Ψ anrichten?
“Modifikationen spielen in allen Aspekten der tRNA-Funktion eine Rolle, von der Struktur und Stabilität bis hin zur Aminoacylierung und Dekodierung am Ribosom. Modifikationen, die die Stabilität oder Faltung der tRNA beeinflussen, treten in der Regel im Hauptteil der tRNA außerhalb der Antikodon-Stamm-Schleife auf. Im Vergleich zu unmodifizierten, in vitro transkribierten tRNAs weisen native, modifizierte tRNAs eine erhöhte Schmelztemperatur, stabilisierte tertiäre Wechselwirkungen und eine erhöhte Gesamtstabilität auf. […] Studien zeigen, dass Pseudouridin die Basenstapel-Wechselwirkungen stabilisiert, während Dihydrouridin im D-Arm die Flexibilität der tRNA bei Organismen erhöht, die bei niedrigen Temperaturen optimal wachsen.”27
“Zahlreiche Untersuchungen haben jedoch gezeigt, dass die Sequenzen im Körper einer tRNA genau auf die Aminoacylierung und die Interaktion mit dem Ribosom abgestimmt sind. Eine Sequenzänderung im tRNA-Körper kann daher zu Unterschieden in der Translationseffizienz führen. Andererseits ist bekannt, dass bestimmte tRNAs an extratranslationalen Funktionen beteiligt sind.”28
Und nun stellen wir uns vor, dass das (m1Ψ) aus der modRNA, aus Versehen, falsch verbaut wird.
Es wird auch vermutet, dass tRNA-Isodekoder regulatorische Funktion beim Housekeeping-Genen spielen. “Ein Housekeeping-Gen (zu Deutsch: Konstitutives Gen oder Haushaltsgen) ist ein Gen, das in allen Zellen eines Organismus dauerhaft und unabhängig von Gewebetyp, Entwicklungsstadium oder äußeren Einflüssen abgelesen (exprimiert) wird.” 29
Die von diesen Genen codierten Proteine und RNAs sind für das nackte Überleben und die grundlegende Aufrechterhaltung der Zelle (den zellulären „Haushalt“) absolut essenziell.
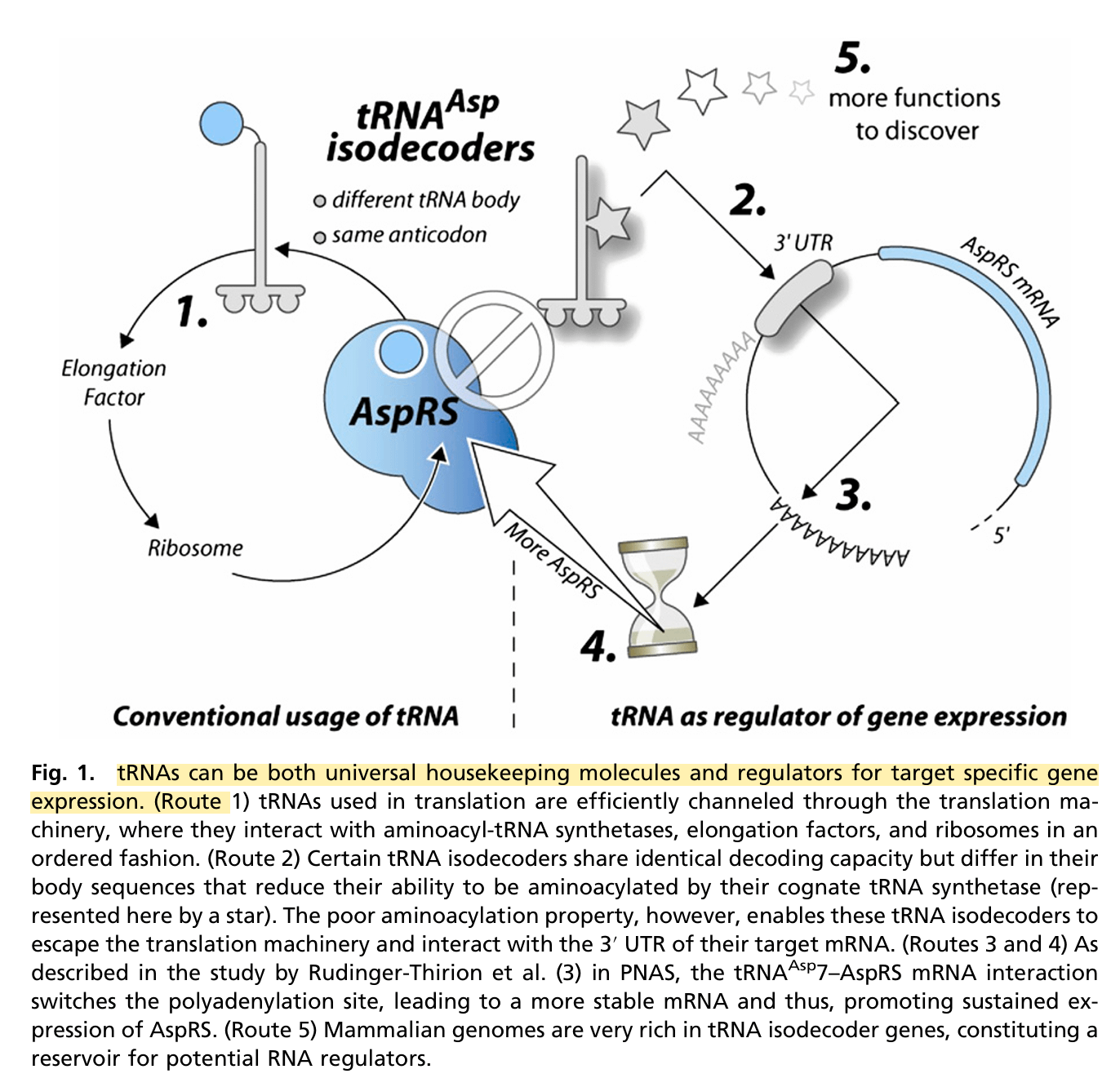
“tRNAs können sowohl universelle „Haushaltsmoleküle“ als auch Regulatoren für die zielspezifische Genexpression sein. (Weg 1) Bei der Translation verwendete tRNAs werden effizient durch den Translationsapparat geleitet, wo sie in geordneter Weise mit Aminoacyl-tRNA-Synthetasen, Elongationsfaktoren und Ribosomen interagieren. (Weg 2) Bestimmte tRNA-Isodecoder verfügen über identische Dekodierungsfähigkeit, unterscheiden sich jedoch in ihren Körpersequenzen, was ihre Fähigkeit zur Aminoacylierung durch ihre kognitive tRNA-Synthetase (hier durch ein Sternchen dargestellt) einschränkt. Die schlechte Aminoacylierungsfähigkeit ermöglicht es diesen tRNA-Isodecodern jedoch, der Translationsmaschinerie zu entkommen und mit der 3′-UTR ihrer Ziel-mRNA zu interagieren. (Wege 3 und 4) Wie in der Studie von Rudinger-Thirion et al. in PNAS beschrieben, verschiebt die Wechselwirkung zwischen tRNAAsp7 und der AspRS-mRNA die Polyadenylierungsstelle, was zu einer stabileren mRNA führt und somit die anhaltende Expression von AspRS fördert. (Weg 5) Säugetiergenome sind sehr reich an tRNA-Isodecoder-Genen, die ein Reservoir für potenzielle RNA-Regulatoren darstellen.30
Kurzum, tRNA-Isodecoder haben noch ein paar Jobs neben der Übersetzung von mRNA in Proteine. Blöd, wenn sie mit der Verbau von (m1Ψ) aus den modRNA-Produkten verbogen sein könnten und ihren Aufgaben nicht nachkommen können.
Zu diesen Nebenjobs/Minijobs gehören:
“Sowohl intakte tRNAs als auch tRNA-Hälften binden an Cytochrom C, wodurch die Aktivierung von Caspasen blockiert und die stressinduzierte Apoptose verringert wird. […] Andere Prozesse, darunter die Modulation der zellulären Reaktion auf DNA-Schäden, werden durch spezifische tRNA-Fragmente reguliert. Die Fähigkeit von tRNA-Fragmenten, die Genexpression zu regulieren, erstreckt sich auch auf die epigenetische Vererbung. tRNA-Fragmente kommen in großen Mengen in den Spermien von Säugetieren vor.”31
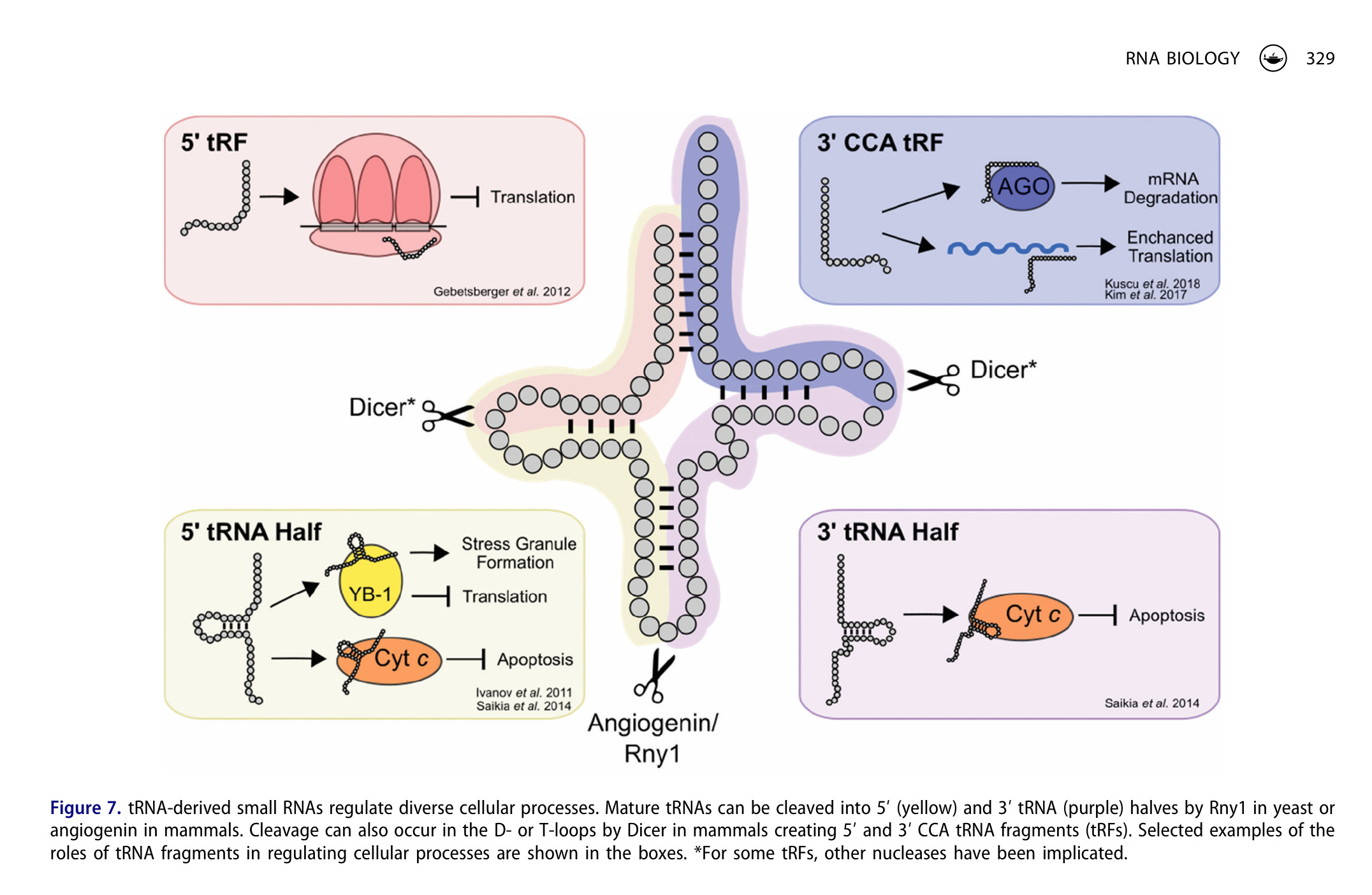
“tRNAs spielen somit eine Rolle bei der extrazellulären Signalübertragung und Regulation. Ein verwandtes Beispiel stammt von Chiou et al., die nachweisen, dass aktivierte T-Zellen tRNA-Fragmente ausscheiden, die die T-Zell-Aktivierung unterdrücken. Extrazelluläre tRNAs und tRNA-Fragmente stoßen auch aufgrund ihres diagnostischen Potenzials auf großes Interesse, da sie in Körperflüssigkeiten nachgewiesen werden. Die genaue RNA-Zusammensetzung in extrazellulären Vesikeln ist vielfältig und umfasst viele Klassen kleiner nichtkodierender RNAs. Interessanterweise stellen Shurtleff et al. fest, dass reife tRNAs in den von HEK293T-Zellen freigesetzten Vesikeln reichlich vorhanden sind. Die Bedeutung extrazellulärer Vesikel bei zellulären Prozessen wie der Reparatur von Muskelfasern und bei Erkrankungen legt nahe, dass extrazelluläre tRNAs eine weitreichende Rolle in der Physiologie von Säugetieren spielen. Angesichts der Bedeutung von RNA-Fragmenten für zahlreiche Aspekte der Genregulation ist es nicht überraschend, dass tRNA-Fragmente eine wesentliche Rolle bei der Entwicklung und bei Erkrankungen spielen. In einer faszinierenden Studie, die auf das therapeutische Potenzial von tRNA-Fragmenten hindeutet, stellen Goodarzi et al. fest, dass tRNA-Fragmente bekannte onkogene Transkripte vom RNA-bindenden Protein YBX1 verdrängen, wodurch deren Stabilität und Expression verringert und infolgedessen das Fortschreiten der Metastasierung unterdrückt wird.”33
“Veränderungen der tRNA-Sequenz und Modifikationen, die mit Krebs, Stoffwechselstörungen und neurodegenerativen Erkrankungen in Verbindung gebracht wurden, sind dokumentiert,”34
“Die Genome von Säugetieren sind sehr reich an tRNA-Isodekodergenen, die potenziell regulatorische Funktionen erfüllen können. So enthält beispielsweise das menschliche Genom 21 Seryl-tRNA-Isodekoder, von denen fünf eine geringe Translationsaktivität aufweisen und vier in vivo bei der Translation vollständig inaktiv sind. Die Identifizierung von tRNA-Regulatoren und deren RNA-, Protein- oder anderen molekularen Zielstrukturen wird eine bislang unerwartete Chance für das postgenomische Zeitalter bieten.”35
“tRNA-Isodekoder können große funktionelle Unterschiede aufweisen, was darauf hindeutet, dass einige tRNA-Isodekoder möglicherweise eine andere Funktion als die Translation erfüllen.”36
RNA, welche Proteine und ganze Organellen regulieren kann und die durch m1Ψ beeinträchtigt sein könnte. Hat das irgendjemand auch nur ansatzweise experimentell geprüft oder ist man einfach davon ausgegangen, dass es schon irgendwie schiefgehen wird?
Unterstützungsmöglichkeiten:
Bücherwunschzettel: https://www.amazon.de/registries/gl/owner-view/30LG3DJ4ET90L?ref_=list_d_gl_lfu_nav
Andere Unterstützungsmöglichkeiten für Holgers und meine Forschung:
Konto für Unterstützung für das Projekt Scan 2000
Dr. Merse DE34 4305 0001 0302 7851 75 Sparkasse Bochum
Horst Reissner: IBAN DE51 4401 0046 0406 4514 67
Dr. S. Stebel: https://ko-fi.com/einmalmitprofisarbeiten
Paypal: Sabine.Stebel@gmx.de
Transfer RNA (TRNA). (n.d.). Genome.gov. https://www.genome.gov/genetics-glossary/Transfer-RNA-tRNA
Abbildung 1 Schülerduden Biologie (2004) S. 210 Translation und das allosterische Dreistellenmodell
Butcher SE, Pyle AM. The molecular interactions that stabilize RNA tertiary structure: RNA motifs, patterns, and networks. Acc Chem Res. 2011 Dec 20;44(12):1302-11. doi: 10.1021/ar200098t. Epub 2011 Sep 7. PMID: 21899297. https://pubmed.ncbi.nlm.nih.gov/21899297/
Berg, M. D., & Brandl, C. J. (2020). Transfer RNAs: diversity in form and function. RNA Biology, 18(3), 316–339. https://doi.org/10.1080/15476286.2020.1809197
Abbildung 1 Schülerduden Biologie (2004) S. 203 Genetischer Code
Mauro VP, Chappell SA. A critical analysis of codon optimization in human therapeutics. Trends Mol Med. 2014 Nov;20(11):604-13. doi: 10.1016/j.molmed.2014.09.003. Epub 2014 Sep 25. PMID: 25263172; PMCID: PMC4253638. https://pubmed.ncbi.nlm.nih.gov/25263172/
Hagenbüchle O, Larson D, Hall GI, Sprague KU. The primary transcription product of a silkworm alanine tRNA gene: identification of in vitro sites of initiation, termination and processing. Cell. 1979 Dec;18(4):1217-29. doi: 10.1016/0092-8674(79)90234-4. PMID: 519766. https://pubmed.ncbi.nlm.nih.gov/519766/
Van Haute L, Powell CA, Minczuk M. Dealing with an Unconventional Genetic Code in Mitochondria: The Biogenesis and Pathogenic Defects of the 5-Formylcytosine Modification in Mitochondrial tRNAMet. Biomolecules. 2017 Mar 2;7(1):24. doi: 10.3390/biom7010024. PMID: 28257121; PMCID: PMC5372736. https://pmc.ncbi.nlm.nih.gov/articles/PMC5372736/
Krüger A, Kovalchuk D, Shiriaev D, Rorbach J. Decoding the Enigma: Translation Termination in Human Mitochondria. Hum Mol Genet. 2024 May 22;33(R1):R42-R46. doi: 10.1093/hmg/ddae032. PMID: 38779770; PMCID: PMC11112381. https://pmc.ncbi.nlm.nih.gov/articles/PMC5372736/
Richter R, Pajak A, Dennerlein S, Rozanska A, Lightowlers RN, Chrzanowska-Lightowlers ZM. Translation termination in human mitochondrial ribosomes. Biochem Soc Trans. 2010 Dec;38(6):1523-6. doi: 10.1042/BST0381523. PMID: 21118119. https://pubmed.ncbi.nlm.nih.gov/21118119/
Knight RD, Landweber LF, Yarus M. How mitochondria redefine the code. J Mol Evol. 2001 Oct-Nov;53(4-5):299-313. doi: 10.1007/s002390010220. PMID: 11675590. https://pubmed.ncbi.nlm.nih.gov/11675590/
Eun, H. (1996). Enzymes and nucleic acids. In Elsevier eBooks (pp. 1–108). https://doi.org/10.1016/b978-012243740-3/50004-1
Mauro VP, Chappell SA. A critical analysis of codon optimization in human therapeutics. Trends Mol Med. 2014 Nov;20(11):604-13. doi: 10.1016/j.molmed.2014.09.003. Epub 2014 Sep 25. PMID: 25263172; PMCID: PMC4253638. https://pubmed.ncbi.nlm.nih.gov/25263172/
Berg, M. D., & Brandl, C. J. (2020). Transfer RNAs: diversity in form and function. RNA Biology, 18(3), 316–339. https://doi.org/10.1080/15476286.2020.1809197
Hughes, L. A., Rudler, D. L., Siira, S. J., McCubbin, T., Raven, S. A., Browne, J. M., Ermer, J. A., Rientjes, J., Rodger, J., Marcellin, E., Rackham, O., & Filipovska, A. (2023). Copy number variation in tRNA isodecoder genes impairs mammalian development and balanced translation. Nature Communications, 14(1), 2210. https://doi.org/10.1038/s41467-023-37843-9
Hughes, L. A., Rudler, D. L., Siira, S. J., McCubbin, T., Raven, S. A., Browne, J. M., Ermer, J. A., Rientjes, J., Rodger, J., Marcellin, E., Rackham, O., & Filipovska, A. (2023). Copy number variation in tRNA isodecoder genes impairs mammalian development and balanced translation. Nature Communications, 14(1), 2210. https://doi.org/10.1038/s41467-023-37843-9
Akeson, S., Kakhaki, P. D., Esfahani, N. G., Reinsch, J. L., Barry, M. L., Zamecnik, M., Tzadikario, T., Abu-Shumays, R. L., Garcia, D. M., Koutmou, K. S., & Jain, M. (2025). tRNA isodecoder analysis using Nanopore ionic current signals and deep learning. bioRxiv (Cold Spring Harbor Laboratory). https://doi.org/10.64898/2025.12.27.696718
Goodenbour JM, Pan T. Diversity of tRNA genes in eukaryotes. Nucleic Acids Res. 2006;34(21):6137-46. doi: 10.1093/nar/gkl725. Epub 2006 Nov 6. PMID: 17088292; PMCID: PMC1693877. https://pubmed.ncbi.nlm.nih.gov/17088292/
Goodenbour JM, Pan T. Diversity of tRNA genes in eukaryotes. Nucleic Acids Res. 2006;34(21):6137-46. doi: 10.1093/nar/gkl725. Epub 2006 Nov 6. PMID: 17088292; PMCID: PMC1693877. https://pubmed.ncbi.nlm.nih.gov/17088292/
Hughes, L. A., Rudler, D. L., Siira, S. J., McCubbin, T., Raven, S. A., Browne, J. M., Ermer, J. A., Rientjes, J., Rodger, J., Marcellin, E., Rackham, O., & Filipovska, A. (2023). Copy number variation in tRNA isodecoder genes impairs mammalian development and balanced translation. Nature Communications, 14(1), 2210. https://doi.org/10.1038/s41467-023-37843-9
Hughes, L. A., Rudler, D. L., Siira, S. J., McCubbin, T., Raven, S. A., Browne, J. M., Ermer, J. A., Rientjes, J., Rodger, J., Marcellin, E., Rackham, O., & Filipovska, A. (2023). Copy number variation in tRNA isodecoder genes impairs mammalian development and balanced translation. Nature Communications, 14(1), 2210. https://doi.org/10.1038/s41467-023-37843-9
Argoudelis AD, Mizsak SA. 1-methylpseudouridine, a metabolite of Streptomyces platensis. J Antibiot (Tokyo). 1976 Aug;29(8):818-23. doi: 10.7164/antibiotics.29.818. PMID: 993120. https://pubmed.ncbi.nlm.nih.gov/993120/
Wurm JP, Griese M, Bahr U, Held M, Heckel A, Karas M, Soppa J, Wöhnert J. Identification of the enzyme responsible for N1-methylation of pseudouridine 54 in archaeal tRNAs. RNA. 2012 Mar;18(3):412-20. doi: 10.1261/rna.028498.111. Epub 2012 Jan 24. PMID: 22274954; PMCID: PMC3285930. https://pmc.ncbi.nlm.nih.gov/articles/PMC3285930/
Halma, M., Raszek, M., & Varon, J. (2025). N1-Methyl-Pseudouridine: The evolution, impact, and future of a key mRNA vaccine modification. Preprints.org. https://doi.org/10.20944/preprints202507.0047.v1
Berg, M. D., & Brandl, C. J. (2020). Transfer RNAs: diversity in form and function. RNA Biology, 18(3), 316–339. https://doi.org/10.1080/15476286.2020.1809197
Berg, M. D., & Brandl, C. J. (2020). Transfer RNAs: diversity in form and function. RNA Biology, 18(3), 316–339. https://doi.org/10.1080/15476286.2020.1809197
Geslain R, Pan T. Functional analysis of human tRNA isodecoders. J Mol Biol. 2010 Feb 26;396(3):821-31. doi: 10.1016/j.jmb.2009.12.018. Epub 2009 Dec 21. PMID: 20026070; PMCID: PMC2822071. https://pubmed.ncbi.nlm.nih.gov/20026070/
Wikipedia-Autoren. (2005, October 27). Haushaltsgen. https://de.wikipedia.org/wiki/Haushaltsgen
Geslain, R., & Pan, T. (2011). tRNA: Vast reservoir of RNA molecules with unexpected regulatory function. Proceedings of the National Academy of Sciences, 108(40), 16489–16490. https://doi.org/10.1073/pnas.1113715108
Berg, M. D., & Brandl, C. J. (2020). Transfer RNAs: diversity in form and function. RNA Biology, 18(3), 316–339. https://doi.org/10.1080/15476286.2020.1809197
Berg, M. D., & Brandl, C. J. (2020). Transfer RNAs: diversity in form and function. RNA Biology, 18(3), 316–339. https://doi.org/10.1080/15476286.2020.1809197
Berg, M. D., & Brandl, C. J. (2020). Transfer RNAs: diversity in form and function. RNA Biology, 18(3), 316–339. https://doi.org/10.1080/15476286.2020.1809197
Akeson, S., Kakhaki, P. D., Esfahani, N. G., Reinsch, J. L., Barry, M. L., Zamecnik, M., Tzadikario, T., Abu-Shumays, R. L., Garcia, D. M., Koutmou, K. S., & Jain, M. (2025). tRNA isodecoder analysis using Nanopore ionic current signals and deep learning. bioRxiv (Cold Spring Harbor Laboratory). https://doi.org/10.64898/2025.12.27.696718
Geslain, R., & Pan, T. (2011). tRNA: Vast reservoir of RNA molecules with unexpected regulatory function. Proceedings of the National Academy of Sciences, 108(40), 16489–16490. https://doi.org/10.1073/pnas.1113715108
Geslain, R., & Pan, T. (2009). Functional analysis of human tRNA isodecoders. Journal of Molecular Biology, 396(3), 821–831. https://doi.org/10.1016/j.jmb.2009.12.018
Tja, dieses elegante einfache Verfahren, wo nach 20 Minuten alles wieder aus dem Gewebe verschwunden ist... Peinlich, dass ich mal in dieser Uni gelernt und unterrichtet habe, wo dieses Wesen jetzt im Vorstand herumgeistert. Aber man kann eben nicht alles haben!
Für die sind auch sämtliche Persistenzrekorde bei Antikörpertitern völlig irrelevant - da sind die Gesichter genauso leer wie bei Tubariusdrüsen, SLYM, Proteinfehlfaltung, Chaperone, reverser Transkription, "Calamari Clots", GMP, GDP, Legaldefinitionen, ICD, ICECI ... von Aaron Siri, vergrabenen Studien, oder selbst EU-geförderterten Nanomedizin-Projektarbeiten ganz zu schweigen.